
3D Printed Models Designed in Blender
All 3D printed models below have been designed in Blender, an open source 3D modeling software, and have been printed on an Elegoo Saturn 3 resin 3D printer. Some of the basic meshes for these models have been used in 3D animations and can been seen on the 3D animations section of this website. The key difference between models for animations and models for 3D printing is that animations can make use of low resolution meshes that have a simple image texture mapped onto them. In contrast, these 3D models for 3D printing require further sculpting because the details need to be a part of the mesh, and that requires a much higher resolution mesh to be able to represent those details. Although these higher resolution 3D models could be used for animations, the time required to render the frames for the animations would take orders of magnitude longer. So, although the models seen in 3D animations and the models seen for 3D printing may appear similar, they were created using very different methods and have very different mesh resolutions. Thanks for reading! -Scott
The heart urchin Echinocardium and its burrow
Sea urchins are echinoderms (echino = spiny; derm = skin) that possess an arrangement of spines around their body (a mineralized skeleton called a test) that can be used for different purposes. These purposes depend upon whether the sea urchin is a regular or irregular sea urchin. Most people are probably familiar with regular sea urchins like those that one might find in tide pools. Regular sea urchins tend to be round with a radial symmetry, meaning that they don’t really have a front or rear. Their mouth is centrally located on their underside, and their anus is centrally located on their top side. They move equally well in all lateral directions as they travel across the top of the seafloor, scraping algae from rocks and clipping kelp. Regular sea urchins do not burrow and are, therefore, exposed to predators at the seafloor surface. Fortunately, regular sea urchins have an arrangement of spines that radiate outward and serve as a form of protection against most predators. One exception to this defense against predators is the sea otter. The sea otter manages to bypass the sea urchin’s defenses by simply cracking the sea urchin’s test open with a rock. Irregular sea urchins do not possess radial symmetry (hence why they are irregular) and do in fact have a front and a rear end that gives them bilateral symmetry. Having a front and a rear means that irregular sea urchins move front-first with their rear trailing behind them. In irregular sea urchins, the mouth is located on the underside of the body but is offset toward the front. The anus is still located on the top side of the body but is offset toward the rear. Unlike regular sea urchins that are confined to the top of the seafloor, irregular sea urchins are capable of burrowing down through the seafloor. Although the spines of a sea urchin may seem like a detriment to burrowing, irregular sea urchins usually have shorter spines than regular sea urchins, and the spines also tend to be swept back across their body toward the rear. This prevents irregular sea urchins from snagging their spines while burrowing. Irregular sea urchins include both the rounder heart urchins and the flatter sand dollars. Herein, we focus on the heart urchin Echinocardium that has been studied extensively for its burrowing behavior. When burrowing, Echinocardium uses shovel-shaped spines on its front and underside to both move and begin to excavate sediment. Once the sediment is loose, spines on the top and sides of the body transport this sediment toward the rear of the sea urchin and compact it against the walls of the burrow behind it. This transportation of sediment from front to rear produces what is referred to as a meniscate backfill. A meniscate backfill, in this case, consists of concave layers of sediment that fill the burrow behind the sea urchin and reflect the shape of the sea urchin’s rear end that is packing sediment behind it. For breathing, feeding, and excreting, Echinocardium uses its spines and tube feet to construct a respiratory channel above their body and a drain for waste removal behind them. Tube feet are small tube-like appendages capable of producing locomotion, grasping, and even respiration. When burrowing below the seafloor, Echinocardium will use its long tube feet to excavate a narrow channel up to the seafloor’s surface, even when its body is sometimes 10-15 cm below the surface. Through this channel, it can use different types of tube feet to breathe, as well as gather food particles from the surface. When Echinocardium excretes waste, it is passed down from the anus to a subanal tuft of spines and tube feet that protrude from the rear end of the sea urchin. These spines and tube feet work to maintain a drain channel behind the sea urchin that is used for flushing waste (like sediment that has been consumed and excreted) away from the active body chamber of the burrow. Trace fossils that consist of a meniscate backfilled burrow with a single drain, like that produced by Echinocardium, are given the name Bichordites. If two drains are present, similar to burrows produced by the sea urchin Brissopsis, the trace fossil is given the name Scolicia. Brissopsis produces two drains because it possesses two separate and distinct subanal tufts of spines and tube feet rather than one, as in Echinocardium. So, one singular tuft produces one drain (Echinocardium and the trace fossil Bichordites), whereas two tufts will produce two drains (Brissopsis and the trace fossil Scolicia). Echinocardium—looking a bit more like a ball of fur than a ball of spines—and seafloor model based on the work of Bromley and Asgaard (1975) [Bulletin of the Geological Society of Denmark, v. 24, p. 261-281] and designed by Scott Mata in Blender v3.6. Echinocardium was printed on an Elegoo Saturn 3 resin printer using Elegoo ABS-Like Resin. The seafloor block model was printed on a Bambu Lab A1 Mini 3D printer using Sunlu PLA filament.










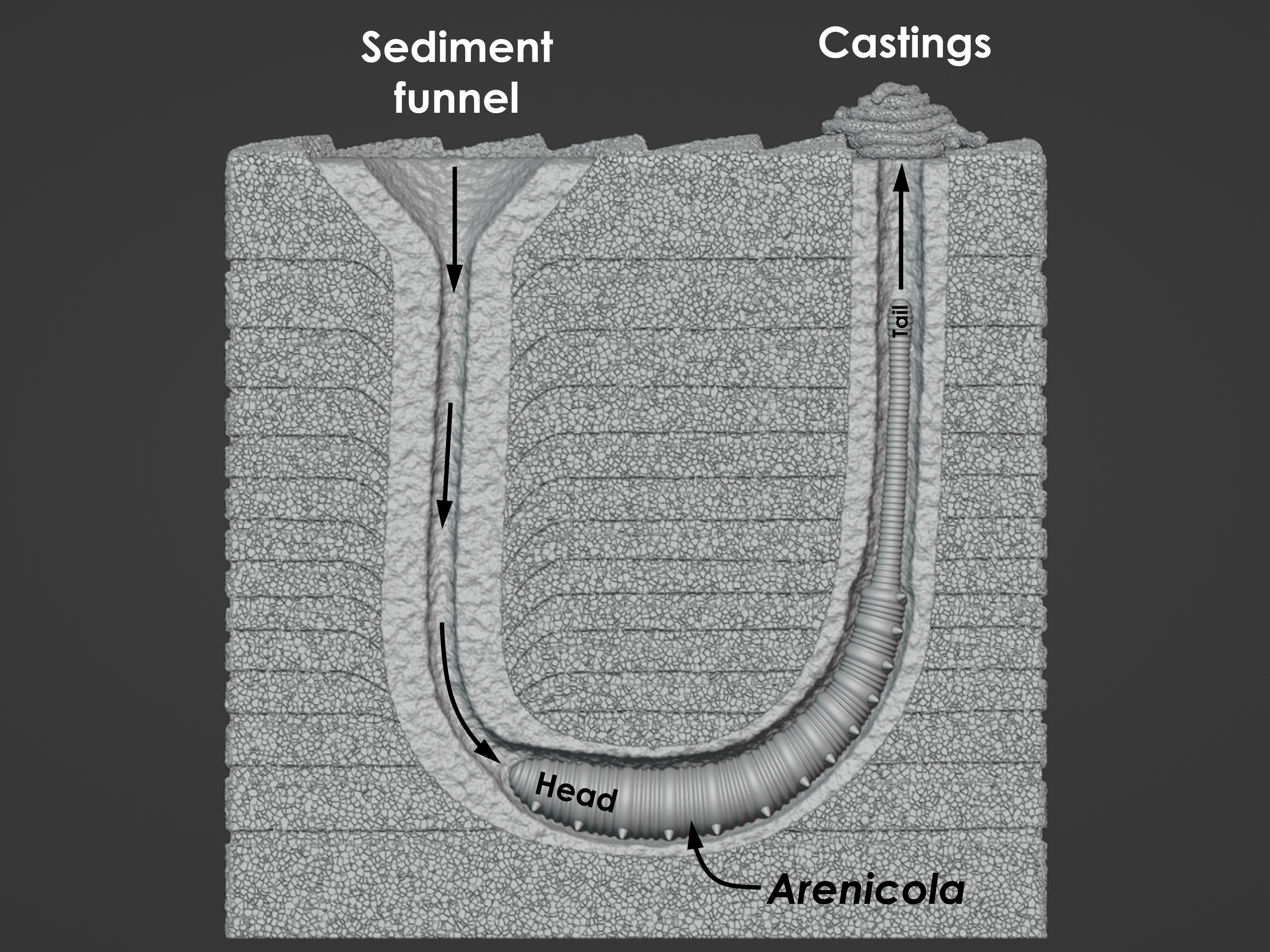
Arenicola and the trace fossil Arenicolites
Annelids are segmented worms that belong to the phylum Annelida. A segmented worm is one that is divided into discrete segments or rings like one might find on an earthworm in a garden. Annelid worms can live on land (example: earthworm), and they can also live in the ocean and along shorelines. Earthworms are, of course, well known for their ability to burrow through dirt and to aerate soils. The same is true for marine annelids that live within seafloor mud or shoreline sand. They like to burrow. One common marine annelid is the lugworm Arenicola that produces very distinctive structures through its burrowing activity. When Arenicola finds itself exposed along the surface of a shoreline, it will begin to burrow into the sediment. As it burrows downward, headfirst, it will bore out an L-shaped or J-shaped burrow. The front half of its body occupies the horizontal base of the burrow, and the tail end of its body occupies the vertical column of the burrow that leads back up to the surface. For Arenicola, this burrow is its home, and it doesn’t ever really need to leave. In the fossil record, there is a trace fossil that goes by the name of Arenicolites. The trace fossil Arenicoltes is defined as a U-shaped burrow, and it is obviously named after the lugworm Arenicola. And yet, Arenicola occupies L-shaped and J-shaped burrows. So, what’s going on? Arenicola may occupy an L- or J-shaped burrow, but its activities within the burrow will actually expand the structure into a U-shape that extends well beyond the body of the worm. Arenicola, of course, needs to feed, but its head is found at the dead end of a burrow below the surface of the seafloor. Fortunately, Arenicola is capable of ingesting sand and mud, which it has in abundance right in front of its head and mouth. Arenicola is a deposit feeder, which means that it ingests large quantities of sediment to extract and process the food contained within it. For sand and mud found along a shoreline, Arenicola finds nutritional value in any bacteria, algae, or other microorganisms that might exist among and between the sediment grains that it consumes. Arenicola’s gut is perfectly capable of extracting nutrients from the organic components of the sediment, but what about all the sand and mud that it can’t digest? For this, Arenicola will extend its tail end up the vertical column of the burrow, toward the surface, and excrete the sediment as castings. Castings consist of long, slender piles of sand and mud deposited at the surface opening of the burrow by Arenicola, and they represent the processed sediment that passed through the gut of the worm, was stripped of its nutrients, and excreted as waste at the surface. This casting pile doesn’t grow infinitely. Rather, it’s usually washed away with the tide or with passing waves. As for the U-shaped nature of the burrow, it's a consequence of this same deposit feeding activity. Arenicola can be thought of as a miner, and its relentless extraction of sediment at depth leads to a systematic mine collapse above its head. As Arenicola consumes, digests, and excretes sediment, it produces a void near its head. This void is not stable, and sediment will collapse straight downward toward the head of Arenicola to fill the void and resupply its food source. This sediment collapse takes the shape of a vertical column or tube with a funnel-shaped depression at the surface, like a tiny sinkhole. This continuously collapsing column combines with the horizontal and vertical sections of the occupied burrow to form the full U-shaped trace fossil that we call Arenicolites. This is not to say that all Arenicolites trace fossils in the rock record are produced by lugworms, or even by this behavior, but this is one well-documented behavior that is known to result in U-shaped trace fossils like Arenicolites. Arenicola (the worm) and seafloor block model designed by Scott Mata in Blender v3.6. Arenicola (the worm) was printed on an Elegoo Saturn 3 resin printer using Elegoo ABS-Like Resin. The seafloor block model was printed on a Bambu Lab A1 Mini 3D printer using Sunlu PLA filament.



Archimedes the corkscrew bryozoan
Bryozoans are a group of marine invertebrates known to build a wide range of mineralized structures and skeletons that can superficially resemble those produced by corals. Despite bryozoans and corals both producing mineralized structures, and both being members of reefs at different points in earth’s history, the two groups really aren’t closely related at all. Corals are cnidarians, with close relatives being animals like sea anemones and jellyfish. Bryozoans are lophophorates, so named because they possess a tentacle-covered feeding organ called a lophophore. The other common animal group with a lophophore would be the brachiopods, and they have their own superficiality with bivalve mollusks like clams. Even though bryozoans form sizeable structures that can be part of even larger reefs, the actual animals living within the structure are in fact much smaller. A single, solitary, bryozoan animal called a zooid is often only about 0.5 mm in size, and a mineralized bryozoan skeleton many house hundreds, if not thousands, of these zooids. When bryozoans are preserved in the fossil record, they often resemble a meshwork filled with hundreds to thousands of holes. This is because the soft-bodied zooids were lost to decay during preservation, leaving behind a mineralized skeleton filled with holes for every single zooid that had lived within the structure. So, the fossil record of bryozoans is one of small and large mineralized skeletal structures that contain hundreds to thousands of small (approximately 0.5 mm) holes representing the former presence of soft-bodied zooids. These structures can take many forms, including domes, branching forms, and flat encrustations on hard surfaces. One unique form belongs to that of the fenestrate bryozoans. Fenestra (as in fenestrate) is Latin for window. Fenestrate bryozoans often resemble thin, fan-like sheets filled with holes (windows) that pass all the way through their structure, producing a mesh-like appearance. One even more unique member of the fenestrate bryozoans was the genus Archimedes that lived during the Carboniferous and Permian Periods (approximately 350 million to 250 million years ago). Archimedes takes its name from the mathematician Archimedes that first described the use of screw pumps in ancient Egypt over 2000 years ago. Screw pumps are used for pumping water, and similar pumps—albeit modernized—are still used today. Archimedes, the bryozoan, takes the form of a screw at its core. Around this core, fan-like sheets of fenestrate mineralized skeleton spiral upward, following the path of the screw. Unfortunately, this type of spiral structure does not preserve well in the fossil record. Rather, what is often preserved is the screw-shaped core in isolation that is far more structurally stable. If the spiraling sheets are preserved in association, they are usually broken off and shattered to pieces, so finding an Archimedes bryozoan in life position, or even whole for that matter, is incredibly rare. The best we can do is reconstruct Archimedes based on the information we have, putting the pieces back together again into a single spiral structure around a screw-shaped core, and that is what is shown in this 3D model below with Archimedes rooted into the seafloor. This 3D model can be repeated upward to just about any height a 3D printer can handle. The major limitations are the tightness of the spiral and the size of the holes. If the spirals are any closer than they currently are in the model, it’s nearly impossible to reach inside to clean, sand, prime, paint, or varnish. If the holes are any smaller, at the current printable scale of the model, it’s nearly impossible to keep them from being clogged by resin, primer, paint, or varnish. So, as a digital 3D model, the spiral could be much tighter, and the holes could be much smaller, but neither would be practical for a model that is going to be 3D printed, cleaned, primed, painted, and varnished. Archimedes 3D model designed by Scott Mata in Blender v3.6 and printed on an Elegoo Saturn 3 resin printer using Elegoo ABS-Like Resin.






Kimberella and the trace fossil Kimberichnus
The Cambrian Period saw the emergence of many modern animal groups. It’s also notable for the widespread adoption of hard, mineralized body parts like shells, bones, and teeth that preserve far better than soft tissues. The time period before the Cambrian is referred to as the Ediacaran Period. Rocks from the Ediacaran Period do preserve animal fossils as well, but it’s in a way that doesn’t rely upon the durability and preservation potential of mineralized parts. This is because Ediacaran organisms, by all accounts, didn’t have hard parts to preserve. Rather, they were soft-bodied and are preserved only as impressions on the seafloor, usually after being rapidly buried. In addition to rapid burial, the presence of microbial mats in the environments where these Ediacaran organisms lived also contributed to their anomalous preservation. Microbial mats in the Ediacaran Period were able to bind sediment of the seafloor together and hold the animal’s form as an impression, allowing for a finer and longer-lasting chance at preservation. It is in this type of microbial mat-dominated marine environment that a soft-bodied animal named Kimberella lived and was preserved. Kimberella is often considered to be a mollusk, similar to a gastropod (example: snail), or is at least considered to be some primitive precursor animal to the group that would later yield mollusks. Kimberella had a body that varied from egg-shaped to pear-shape in outline, which might imply that the body was fully capable of stretching along its length. It had a shell-like covering on its back that was likely soft, rather than mineralized, due to observed elongation and deformation during preservation. Kimberella had a wavy fringe, similar to a pleated skirt, that encircled most of its perimeter and extended outward from what might be described as a muscular foot similar to that of a gastropod like a snail. The muscular foot of a gastropod is the part of the body that contacts the ground and is responsible for locomotion. At what appears to be the front of the animal is a head with a feeding organ lined with what would have possibly been teeth, although teeth aren’t directly preserved. The reason these teeth are considered to have existed is because Kimberella fossil impressions are associated with a trace fossil referred to as Kimberichnus that reflects the use of these supposed teeth. The trace fossil Kimberichnus consists of a set of linear grooves arranged into a fan shape, and these traces are often found associated with microbial mat features and Kimberella impressions in the rock record during the Ediacaran Period. The trace fossil Kimberichnus is interpreted as the result of Kimberella using an arc-shaped set of teeth on its feeding organ to rake through a microbial mat. This raking motion would leave behind a set of linear grooves, corresponding to the arrangement of the teeth, as the microbial mat is ripped up from the seafloor for consumption by Kimberella. The Kimberella body and Kimberichnus trace fossil 3D models are based on research and ecological interpretations by Ivantsov (2009) [Paleontological Journal, v. 43, p. 601–611] with some features of Kimberella drawn from the work and interpretations of Fedonkin and Waggoner (1997) [Nature, v. 388, p. 868–871]. Any errors or misinterpretations of anatomy are my own. Kimberella and Kimberichnus 3D models designed by Scott Mata in Blender v3.6 and printed on an Elegoo Saturn 3 resin printer using Elegoo ABS-Like Resin.


















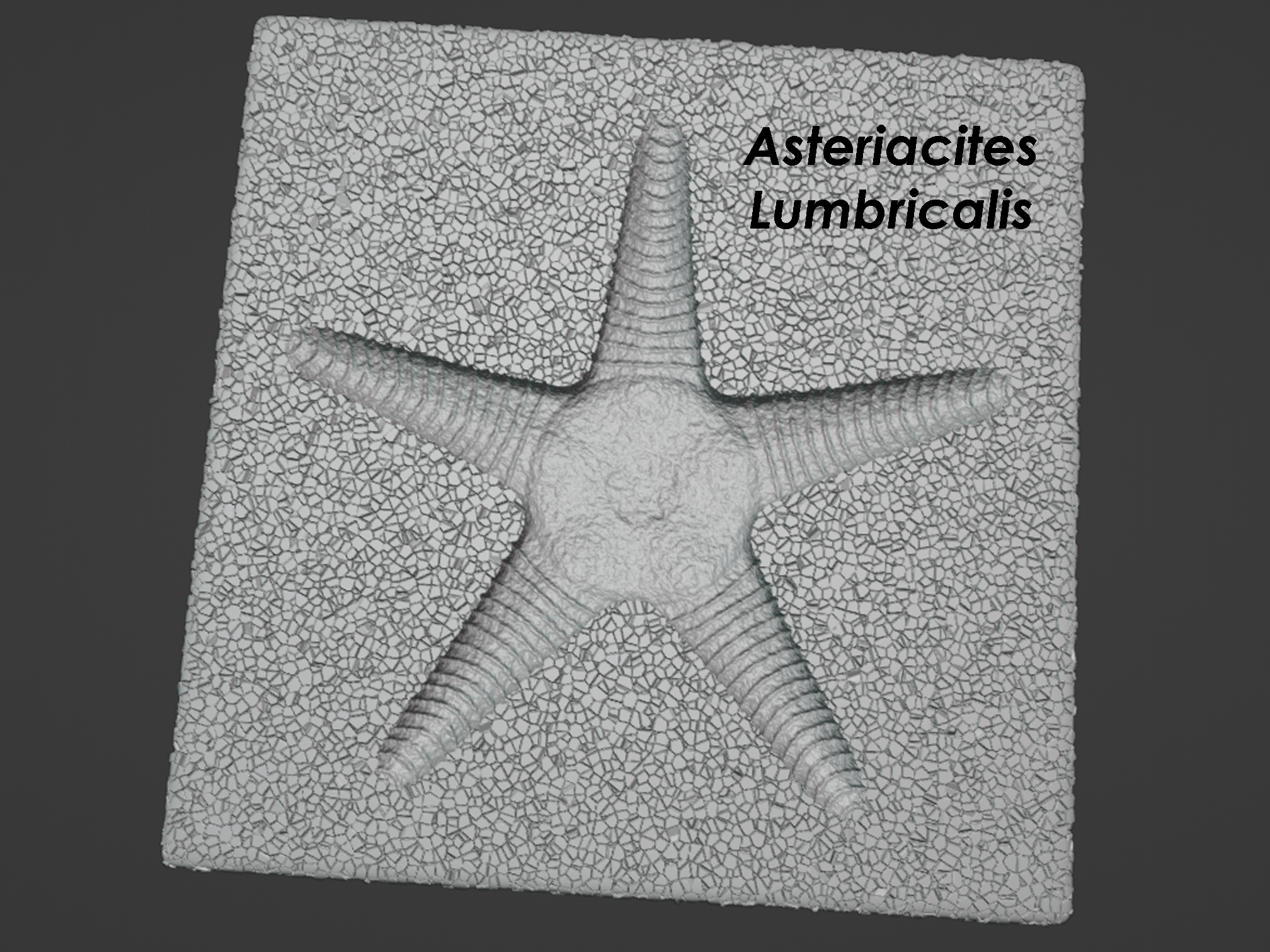
Brittle stars, sea stars, and the trace fossil Asteriacites
Trace fossils represent fossilized behavior. These behaviors may be preserved in many forms, including common examples like burrows, impressions, trackways, and trails. Much like organisms and fossils can be classified by binomial nomenclature using a genus and a species name, so too can trace fossils. Modern humans are classified as Homo sapiens (genus = Homo; species = sapiens). Where trace fossils differ is that they are classified according to their ichnogenus and ichnospecies because they don’t represent actual organisms. Ichnogenus and ichnospecies only represent behaviors, and there are no broader categories like kingdom, phylum, class, order, or family like we would use for organisms themselves. The oldest valid ichnogenus name is Asteriacites, which was described by E.F. von Schlotheim in 1820. Asteriacites represents a star-shaped impression attributed to a star-shaped organism. In the modern ocean, similar star-shaped impressions can be made by sea stars (starfish) and brittle stars, and ancient examples of the trace fossil Asteriacities are attributed to the resting or burrowing behavior of these two organisms. Sea stars and brittle stars both belong to the subphylum Asterozoa (astero = star; zoa = animal) within the larger phylum of echinodermata (echino = spiny; derm = skin). Echinoderms as a phylum would include sea urchins, sand dollars, sea cucumbers, crinoids and, of course, sea stars and brittle stars. Within the ichnogenus Asteriacites are defined three ichnospecies: Asteriacites lumbricalis, Asteriacites stelliformis, and Asteriacites quinquefolius. Asteriacites lumbricalis is attributed to the burrowing or feeding activity of a brittle star as it buried itself in the seafloor. Brittle stars consist of a central disc-shaped body with five delicate arms that radiate out from this body. This is reflected in the trace fossil Asteriacites lumbricalis, and one of the defining characteristics of this ichnospecies is a distinct central disc impression from which radiates the impressions of five slender arms. Asteriacites stelliformis and Asteriacites quinquefolius lack this central disc impression and consist simply of five radiating arms that meet in the middle. Asteriacites stelliformis and Asteriacites quinquefolius are distinguished from each other based upon the relationship between arm length and arm width. Asteriacites stelliformis has an arm length:width ratio greater than 2:1, which would make it look roughly like the shape of a sea star. Asteriacites quinquefolius has an arm length:width ratio less than 2:1, which would make it look less like a star and more like a pentagon. Asteriacites stelliformis can be made by sea stars and would largely reflect the shape of the sea star’s body. Asteriacites stelliformis can also be made by brittle stars in situations where the brittle star sweeps its arms through the sediment to create an impression far more expansive than its central disc and slender arms would normally produce. This behavior would be similar to how humans can sweep their arms and legs through the snow to produce a snow angel impression far more expansive than a typical human silhouette. Asteriacites quinquefolius is usually attributable to only sea stars and reflects a shallow burial in loose sediment with extensive surface disruption that would produce poorly defined arms and a poorly defined shape in general. In one of the models below, the brittle star Ophiolepis superba, also known as the banded serpent star, is shown producing Asteriacites lumbricalis. As is a fairly common posture for brittle stars, the banded serpent star is shown with its mouth facing downward into the sediment while its arms are raised high up into the water above its body. This is relevant because it illustrates that Asteriacites lumbricalis doesn’t necessarily record the full length of the brittle star’s arms. The length of brittle star arms can vary anywhere between 2 to 20 times the diameter of their central disc, yet most fossils of Asteriacites lumbricalis don’t record such extreme arm lengths, likely because a significant portion of the brittle star’s arms were actually floating in the water during burrowing. In the other model below, the sea star Astropecten articulatus, also known as the royal starfish, is shown producing Asteriacites stelliformis. In this instance, the trace fossil is nearly identical in shape and size to the body of the sea star, and all parts of its body and arms are submerged in the sediment and reflected directly in the trace fossil. Brittle star, sea star, and trace fossil block models designed by Scott Mata in Blender v3.6.













Inarticulate brachiopod “burrowing adventure!” playset
Admittedly, it’s a very tongue-in-cheek title. :)
Among animals, the phylum lophophorata had particular prominence in Earth’s ancient oceans (primarily during the Paleozoic Era) but has since become far less common, especially in the modern ocean. Members of the phylum are referred to as lophophorates because they use a tentacle-covered feeding appendage called a “lophophore”. Among the lophophorates are the brachiopods, and these brachiopods can, at times, superficially resemble bivalve mollusks like clams, oysters, and mussels. This superficial resemblance stems from the fact that both groups of animals, bivalves and brachiopods, have a two-part (two-valve) shell that typically hinges and can swing open and closed as needed for feeding and breathing. Brachiopods are considered either articulate (articulated = hinged) or inarticulate. Articulate brachiopods have a two-part shell that is capable of swinging open and closed along a hinge line. This is similar to how bivalves operate. In contrast, inarticulate brachiopods have no hinge at all and are simply not capable of swinging their shell open or closed in that way. Rather, inarticulate brachiopods use muscles to hold the two halves of their shell together without any meaningful hinged connection. What inarticulate brachiopods lack in articulation, they make up for in their ability to burrow. Both articulate and inarticulate brachiopods have a long external (outside the shell) fleshy extension of their body that is called a pedicle. In articulate brachiopods, the pedicle is used to anchor the shell to firm or hard surfaces at the seafloor’s surface. After the shell is anchored, an articulate brachiopod tends to live a stationary existence and doesn’t typically doesn’t change locations, at least not willingly. For inarticulate brachiopods, the pedicle is used for burrowing and mobility. An inarticulate brachiopod can “walk” across the seafloor using the front of its shell and the end of its pedicle in a sort of arched, worm-like motion. The pedicle is also used for anchoring and adjusting an inarticulate brachiopod within a subsurface burrow. Inarticulate brachiopods are known to burrow down into the sediment by leading with the front edge of their shell as their pedicle trails behind them. They accomplish this by using a scissor-like motion of the two valves of their shell to sweep and transport sediment away from their leading edge. Following a u-shaped trajectory, the brachiopod will burrow downward before emerging back upward near the seafloor surface with its pedicle pointing down and the front edge of the shell facing up. In this position, the pedicle serves as an anchor and can push up or pull down the brachiopod’s shell and body to keep it at the seafloor’s surface for feeding. These up and down adjustments of the shell and body are often recorded as downward deflected sediment layers within the burrow, just beneath the brachiopod’s shell. The deflected layers themselves reflect a combination of sedimentation within the burrow, filling-in beneath the shell, as well as actual physical disruption due to up and down shell movement. The inarticulate brachiopod burrowing behavior and seafloor block model shown below were heavily inspired by Zonneveld et al. (2007) [PALAIOS, v. 22, p. 74-97], which was supported by previous work by Thayer and Steele-Petrovic (1975) [Lethaia, v. 8. P. 209-221] and Emig et al. (1978) [Geobios, v. 11, p. 573-609]. The 3D model demonstrates some of the common burrowing styles of inarticulate brachiopods, as well as the movement of an inarticulate brachiopod across the seafloor before beginning its burrowing descent. Inarticulate brachiopods and seafloor block model designed by Scott Mata in Blender v3.6. Inarticulate brachiopods were printed on an Elegoo Saturn 3 resin printer using Elegoo ABS-Like Resin. The seafloor block model was printed on a Bambu Lab A1 Mini 3D printer using Sunlu PLA filament.






Infaunal and semi-infaunal bivalves
Bivalves are mollusks that have a shell arranged into two halves or valves [bi = two; valve = 1 part of a 2-part shell] that hinge together and are capable of opening and closing through muscle movements. Bivalves would include common mollusks like clams, mussels, oysters, and scallops. Among these bivalves, different lifestyles can be recognized. Epifaunal bivalves would be those that live on top of the seafloor or on top of firm or hard substrates like rocks or even the shells and skeletons of other organisms. Mussels would be a good example of an epifaunal bivalve. Mussels will attach themselves to rocks using byssal threads (collectively known as a byssus), which are sticky fibers that utilize a strong biological produced glue capable of adhering the mussel to just about any solid object that they encounter. Infaunal bivalves would be those that live fully submerged (or nearly so) below the seafloor’s surface. Many clams fall into this category. To get below the seafloor, these bivalves need to burrow, much in the same way that a worm would need to burrow to get below the surface. Fortunately, clams have a large muscular foot and a siphon to aid them in this process. A muscular foot is a feature present in nearly all mollusks, although its shape and purpose vary widely. In cephalopods like squid, the muscular foot takes the form of arms and tentacles. In gastropods like snails, the muscular foot is the lower portion of their body responsible for mobility along the seafloor or up a brick wall in a garden. In clams, the muscular foot is an extension of the soft body of the clam that serves as both spade and anchor. For a gardener, a spade is the flattened form of a shovel that is used for cutting down through sediment. The same is true for clams. When the clam wants to burrow down through the seafloor, it will shape its muscular foot into a spade and dig down through the sediment. At depth, the lower end of the foot will expand into a bulbous anchor that will lock itself into place within the sediment. Once secured, the clam will pull itself down into the seafloor by using the muscular foot as a stationary anchor. See the “Burrowing behavior in bivalves” video on the 3D animation section of this website for a more detailed explanation and visualization. Once sufficiently buried below the seafloor, the clam will then send its siphons up to the surface so that it can maintain access to fresh seawater and the oxygen and food contained within it. Siphons are tubes, usually paired, that extend upward from the soft body of the clam. One siphon tube is used for inhaling water: the inhalant siphon. The other is used for exhaling water: the exhalant siphon. After water enters through the inhalant siphon, it passes over the gills where oxygen and food are extracted. Past the gills is the mouth where the food is then consumed. Once oxygen and food are removed, the water is then expelled through the exhalant siphon. This is how a burrowing, infaunal bivalve survives despite its shell and body being largely removed from the seafloor surface. This infaunal lifestyle is illustrated by the clams Mya arenaria and Mercenaria mercenaria in the 3D model shown below. Mya arenaria is shown within a burrow that has been repeatedly deepened over time due to erosion at the surface. This is reflected in the concave-downward laminae in the upper half of the burrow that would have formed one after the other as the bivalve shifted downward, creating a void above its shell to be filled by sediment. Mercenaria mercenaria is shown within a burrow that has been repeatedly shallowed over time due to deposition at the surface. Sediment would have filled-in beneath the clam each time it raised its shell upward, creating a void. Between epifaunal and infaunal is an additional lifestyle called a semi-infaunal lifestyle. Semi-infaunal bivalves live only partially submerged below the seafloor. This is a lifestyle common among pen shells. Pen shells are named as such because they resemble a quill pen with a narrow, pointed base and a flared top. Pen shells average 15-30 cm (6-12 inches) in length but can grow much longer, so one might consider that whoever coined the term pen shell was imagining something large like a goose feather quill with all its barbs intact. Pen shells usually bury a little less than half their shell below the seafloor with the pointed end down and the flared end up. In addition to staking its pointed end within the seafloor for anchorage, the pen shell also makes use of byssal threads. Unlike mussels that attach their byssal threads to rocks above the seafloor, the pen shell will attach its byssal threads to sand grains, gravel, and shell fragments below the seafloor. An adult mussel may use 100 or fewer byssal threads to attach itself to a rock. In contrast, an adult pen shell uses thousands of byssal threads at a time, which would allow it to attach itself to thousands of sediment grains within the soft seafloor. So, although the pen shell lacks what one might consider to be a solid substrate like a rock, it more than makes up for it in sheer volume of attachment points between byssal threads and sediment grains. This semi-infaunal lifestyle is illustrated by the pen shell Atrina pectinata in the 3D model shown below. Bivalves and seafloor block model designed by Scott Mata in Blender v3.6. Bivalves were printed on an Elegoo Saturn 3 resin printer using Elegoo ABS-Like Resin. The seafloor block model was printed on a Bambu Lab A1 Mini 3D printer using Sunlu PLA filament.




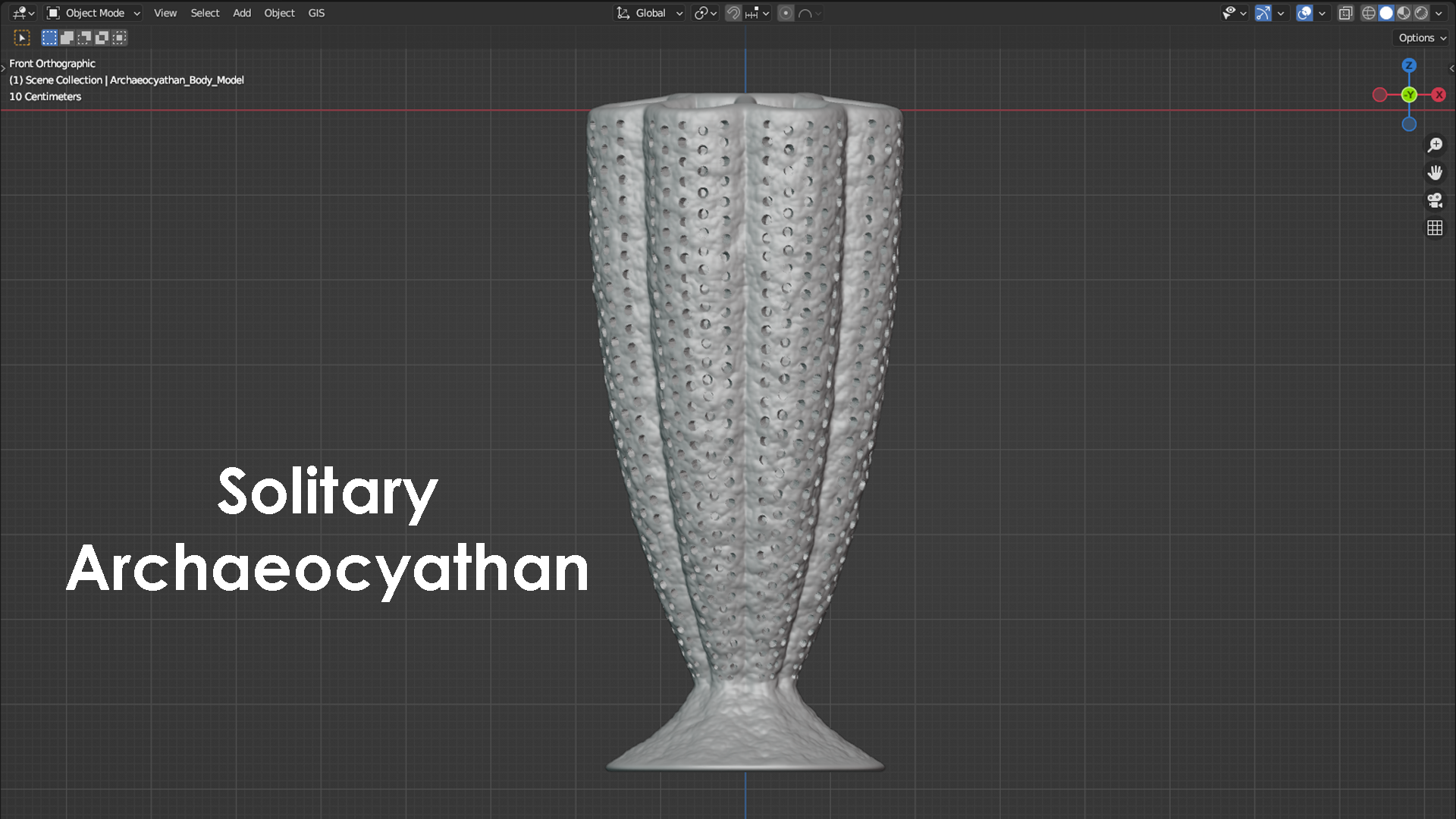
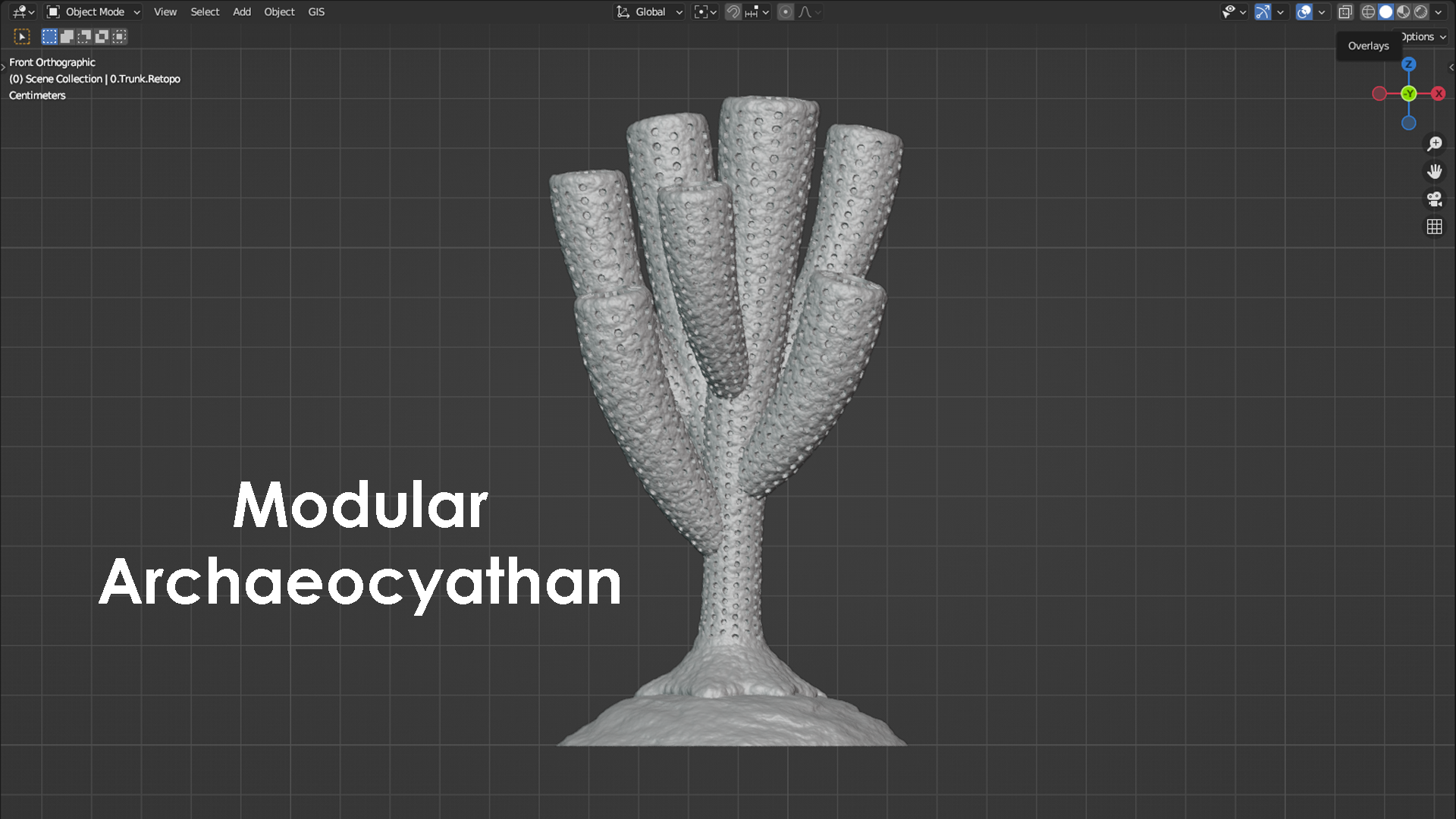
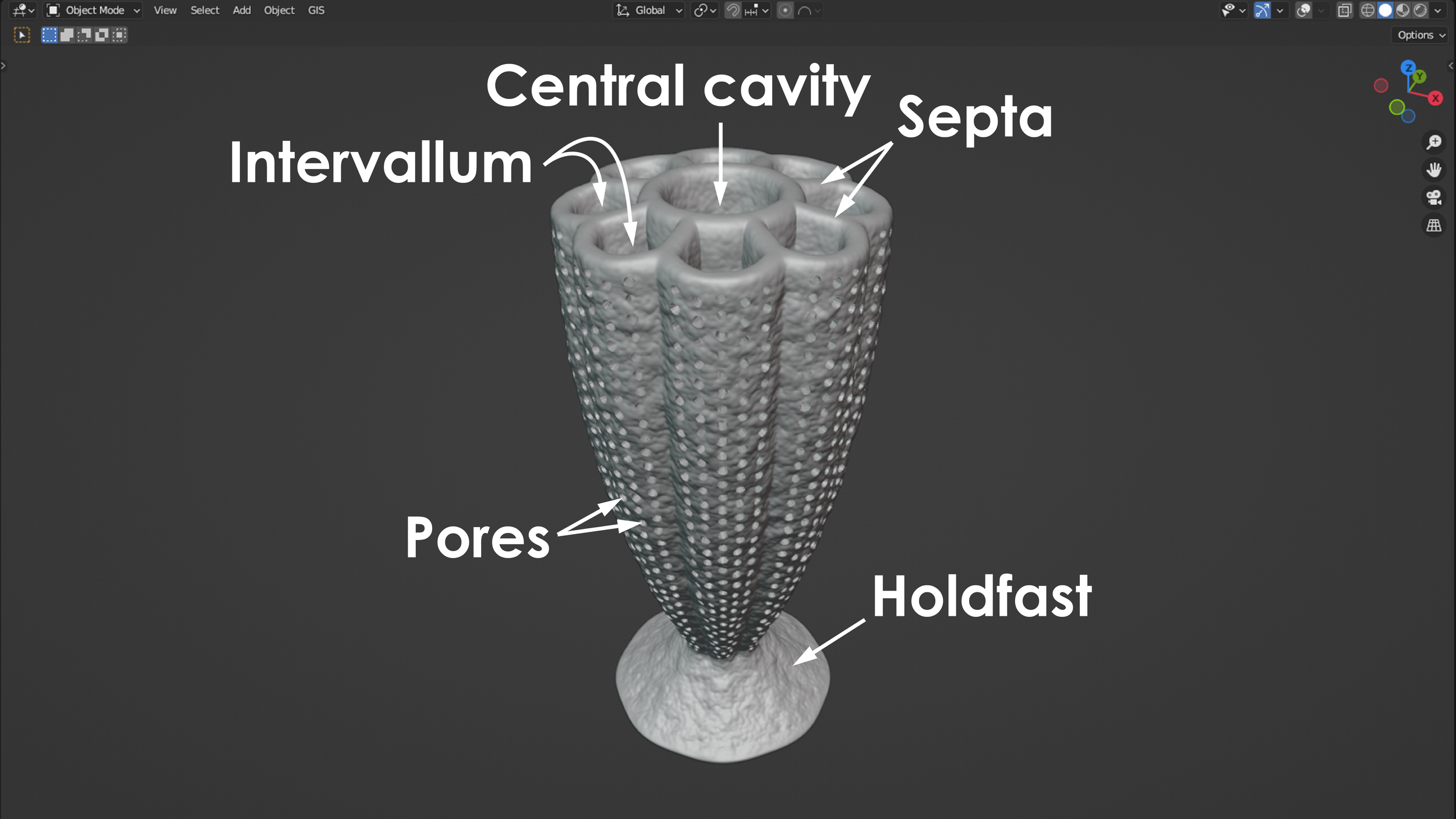
Archaeocyathans and their morphology
The phylum porifera contains all the sponges that exist today. This includes sponges that live in marine environments and those that live in freshwater environments. Porifera also contains many extinct groups and forms that have existed and since perished over the past 600 or so million years. Sponges are animals that contain a system of pores and canals that allow them to filter water through their bodies and extract food particles from that flowing water. This filtering is how they feed, and many sponges have a shape similar to a chimney to filter this water efficiently. Typically, water is sucked in through the outside wall of the sponge, food is filtered and extracted from the flowing water, and the filtered water is then expelled upward through a vertically oriented, central cavity. During the Cambrian Period (approximately 538-485 million years ago), the most common and most diverse members of the porifera phylum were the archaeocyathans. Some interpretations place archaeocyathans as their own phylum apart from the sponges and the phylum porifera, but modern convention considers them as a taxonomic class within the phylum of porifera. Being a member of porifera would suggest that archaeocyathans share many anatomical characteristics with a typical sponge, and they do. Where archaeocyathans differ from a typical sponge is in the structure of their walls. A typical marine sponge has only a single wall between the outside environment and its central cavity. This means that a typical sponge will filter water through only this single wall before evacuating the filtered water upward and outward into the environment. Archaeocyathans, in contrast, are defined by the presence of two walls (inner and outer) that are arranged into what is often referred to as a nested cone-in-cone structure. One might imagine two tapered ice cream cones nested within each other with a vacant space in between, and this would not be too far off from a typical archaeocyathan’s general anatomy. The space between the outer wall and inner wall of an archaeocyathan is called the intervallum, and this corridor is typically divided into compartments separated by additional walls called septa (plural = septa; singular = septum) that span the gap between the inner wall and outer wall. The presence of two distinct walls, an intervallum, and septa are the key characteristics that are present in all archaeocyathans but are lacking in all other members of the phylum porifera. This is what makes archaeocyathans unique among all other sponges. Beyond their distinctive anatomy, archaeocyathans are also notable for emerging, living, thriving, and then going extinct all within the Cambrian Period. During this time interval, archaeocyathans claimed the title of the first animal reef builders and were responsible for creating a rigid framework within the reefs they built and occupied. This is in contrast to modern reefs where the rigid framework is often produced through a combination of growth primarily from coral and algae. Although archaeocyathans produced many different morphologies, two of the most general forms are referred to as solitary and modular (branching). A solitary archaeocyathan consists of a single cone-in-cone structure attached to the seafloor with some sort of holdfast or anchor. Modular archaeocyathans consist of two or more cone-in-cone structures that branch off from a single, central holdfast or anchor. In this regard, modular archaeocyathans can resemble branching colonial corals in their broad form, although neither would really be mistaken for the other upon close inspection. The 3D models shown below represent a very generalized version of a solitary archaeocyathan and of a modular archaeocyathan. The solitary archaeocyathan was inspired by Archaeocyathellus from the Treatise on Invertebrate Paleontology Part E [Fig. 3, p. E4]. The modular archaeocyathan was inspired by the framework-building, branching archaeocyathans documented within the Poleta Formation of Nevada by Rowland (1984) [Geology, v. 12, p. 181-183]. Archaeocyathan models designed by Scott Mata in Blender v3.6 and printed on a Bambu Lab A1 Mini 3D printer using Sunlu PLA filament.



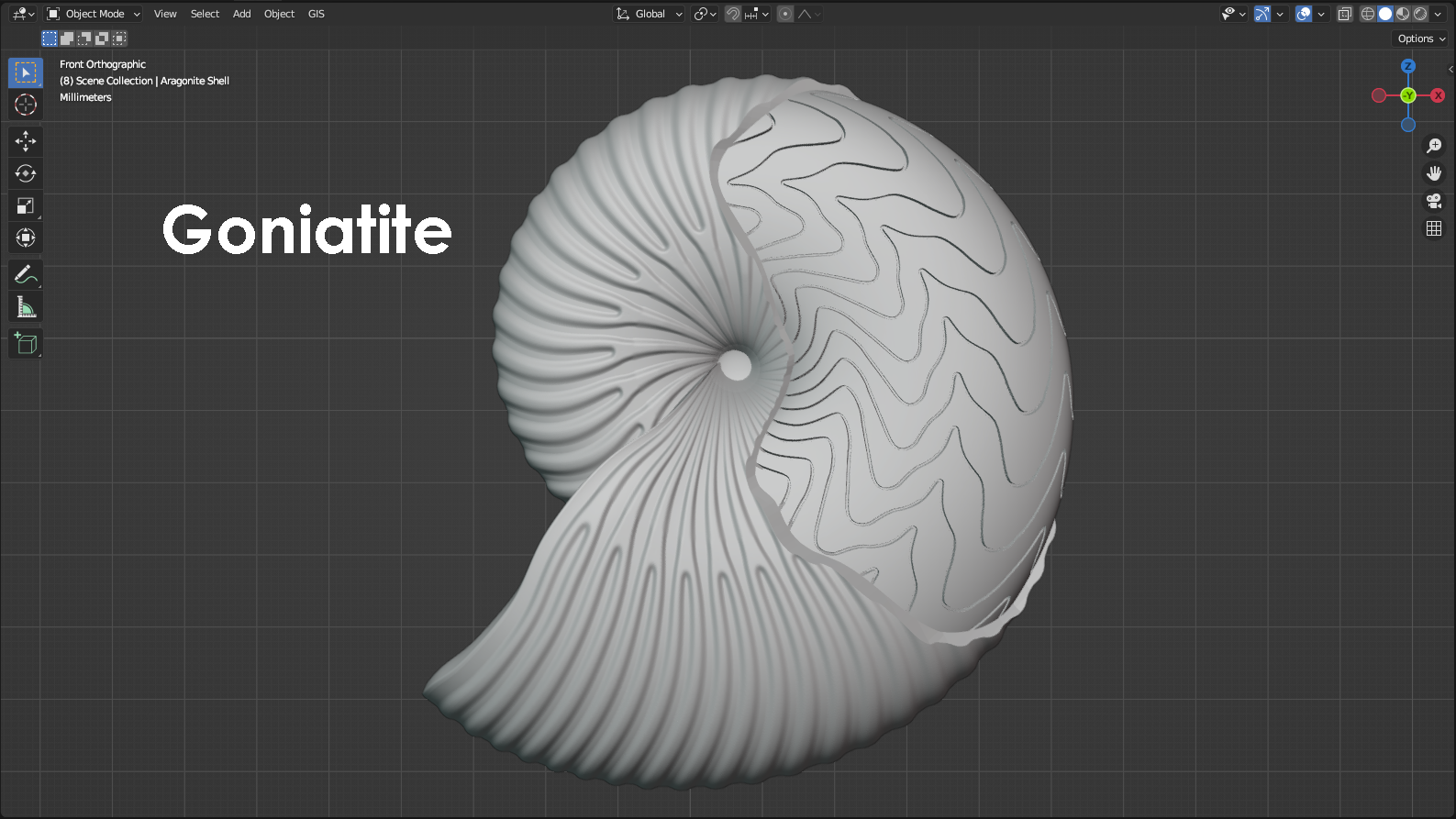
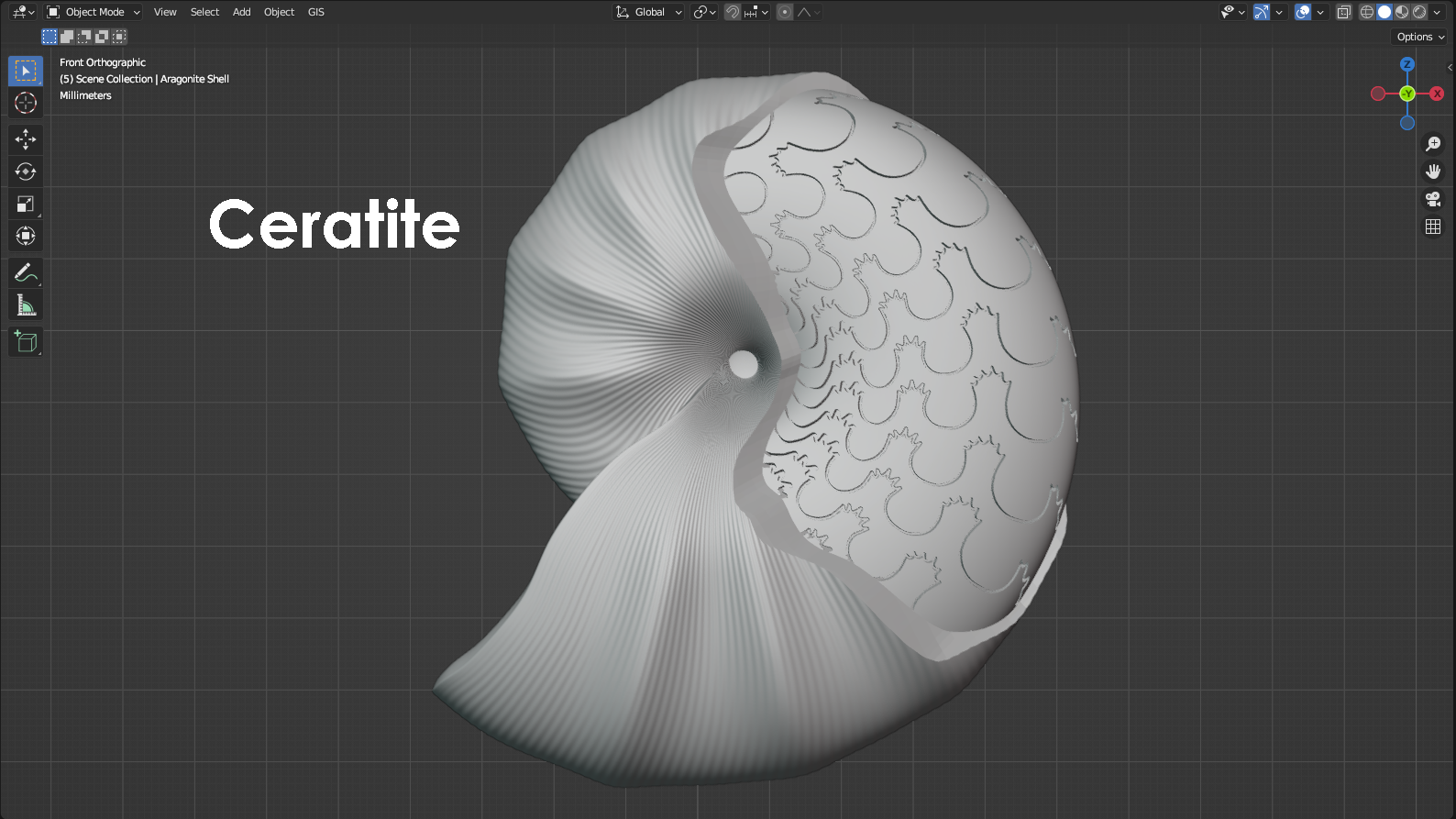
Ammonoids and their suture patterns
Cephalopods are mollusks that have a body organized into a head region that extends down past the mouth into a set of arms and sometimes tentacles. Modern day cephalopods include animals like octopus, squid, cuttlefish, and the chambered Nautilus. Of these listed cephalopods, one is not like the others. That would be the Nautilus. The chambered Nautilus is the only modern cephalopod that has an external mineralized shell. For the Nautilus, the shell is divided into chambers that can be filled with gas and regulated by the Nautilus to adjust the animal’s buoyancy. Adding gas to the internal chambers increases the shell’s buoyancy and allows the Nautilus to rise through the water column, whereas removing gas from the chambers decreases the shell’s buoyancy and allows the Nautilus to sink. Although the presence of an external shell is rare among modern-day cephalopods, that wasn’t the case in the past. In fact, shelled cephalopods are known from up to nearly 500 million years ago, and they only really went into dramatic decline during the Cretaceous-Paleogene mass extinction about 66 million years ago. This is the mass extinction that wiped out the dinosaurs, and it also eliminated many groups of shelled cephalopods. One major group of shelled cephalopods that failed to survive into the modern age is the ammonoids, which includes several different orders. Most notable among them are the goniatites, ceratites, and ammonites. These different orders of ammonoids are distinguished primarily by their “suture patterns” that represent the shape of the walls between chambers inside the shell. When a cephalopod shell is buried within the seafloor, the chambers can fill with sediment or precipitated minerals. Over time, the shell itself can actually dissolve away, leaving behind only the sediment or minerals that filled the interior of the shell. Where the walls between chambers were once located are now recesses that we refer to as “suture patterns” that preserve the shape of the walls, even though the walls are long gone. In the case of the modern Nautilus, the walls between chambers are smooth and gently curved. In the past, with the ammonoids, this wasn’t the case. For the goniatites, the walls between chambers would have been very wavy and, at times, pointed. The smoother, rounded parts of a suture pattern that are convex toward the shell’s opening are referred to as “saddles”, whereas the pointier parts of a suture pattern that are convex away from the shell’s opening are referred to as “lobes”. For the goniatites, the saddles are gently curved, and the lobes are simple, smooth points. For the Ceratites, the saddles are still gently curved, but the lobes consist of hand-like protrusions with small fingers that point away from the shell’s opening. For ammonites, both the saddles and the lobes are complex and contain branching and fractal-like patterns. In terms of complexity, goniatites are considered to have the simplest suture patterns, whereas ammonites are considered to have the most complex suture patterns. Goniatites went extinct during the Permian-Triassic mass extinction (approximately 252 million years ago). Ceratites went extinct during the Triassic-Jurassic mass extinction (approximately 201 million years ago). Ammonites went extinct during the Cretaceous-Paleogene mass extinction (approximately 66 million years ago). The 3D models represented below illustrate these differences between the goniatites, ceratites, and ammonites. Each model has a section of the shell’s outer wall sliced away to reveal what the suture pattern might look like if the ammonoid was preserved as a fossil. For the animal living within the shell, a model was constructed that resembles the anatomy of a modern-day squid with eight arms and two tentacles. A squid tentacle is longer than an arm and terminates in a wide, paddle-like lobe called a “tentacular club”. Tentacles are typically used for ensnaring prey that might be passing by, whereas the arms are used for the more nuanced manipulation of objects. Ammonoid models designed by Scott Mata in Blender v3.6 and printed on an Elegoo Saturn 3 resin printer. Ammonoid shells are printed using Elegoo ABS-Like Resin. The animal itself is printed separately using Elegoo ABS-Like Resin. Although modeled facing left, the ammonoids were mirrored and 3D printed facing right to better show their chronological appearance in the fossil record from goniatites to ceratites to ammonites.


Echinoderm lifestyles during the Cambrian substrate revolution
This set of 3D printed models represents three varieties of echinoderms that lived during the early Cambrian Period and reflect lifestyles that became scarce following the Cambrian substrate revolution as per Bottjer et al. (2000) [GSA Today, v. 10, p. 1-7]. As is described on the 3D animations section of this website for the videos on “Helicoplacoids and the Cambrian seafloor”, “Edrioasteroids and the Cambrian substrate revolution”, and “Adaptation and attachment in early Paleozoic crinoids and eocrinoids”, the Cambrian Period is a time of transition for the seafloor. With the radiation of burrowing organisms, there was a fundamental shift from firm microbial mat-covered seafloors to thoroughly burrowed, water-saturated, soupy seafloors. This shift was problematic for many organisms that had evolved and had built an ecological lifestyle around embedding themselves in a firm seafloor, similar to how a golf tee might embed itself within firm grass. This firmness of the seafloor was a consequence of a general lack of deep burrowing and was, of course, aided by the presence of microbial mats in certain environments to further bind the seafloor. These three echinoderms—Camptostroma, Helicoplacus, and Lichenoides—reflect this lifestyle that is referred to as a sediment sticker (for sticking in firm sediment) or mat sticker (for sticking in microbial mat-bound sediment). Camptostroma is an edrioasteroid that would have lived embedded within a firm seafloor as a sediment sticker or a mat sticker with its pointed lower end stuck straight down into the seafloor. Following the Cambrian substrate revolution, edrioasteroids would adapt to the changing seafloor by flattening and becoming encrusters that would glue their suction cup-like bodies to hard surfaces like the shells of other organisms. Helicoplacus is a helicoplacoid, which is a group of echinoderms known only from the Cambrian Period. Unlike other echinoderms that are known for their five-fold symmetry and can be divided five ways like a sea star, helicoplacoids have three-fold symmetry that can be seen in the three branches of plates that radiate out from the center of their body. Helicoplacoids would have been sediment stickers or mat stickers similar to Camptostroma, and their narrow end would have been embedded into a firm seafloor so that they could stand upright. Following the Cambrian substrate revolution, helicoplacoids seemingly couldn’t adapt to the new softer seafloor and simply went extinct as a group. Lichenoides is an eocrinoid that had a bulbous body divided into somewhat large plates relative to its body size, and these plates are connected with equally large sutures. The lower end of Lichenoides is very round, as compared to Camptostroma and Helicoplacus, and it has 15 arms that radiate outward from the top of its body. Later eocrinoids developed stems and holdfasts that would help them to either root themselves into a soft seafloor or affix themselves to hard surfaces like the shells of other organisms. Lichenoides has no stem and no holdfast, and it’s therefore likely that it rested directly on the seafloor with no particular method of attachment other than possibly embedding itself within a firm seafloor. All three echinoderm models are resting on square bases that allow them to sit upright in their life position. These bases represent the nature of the seafloor that these organisms would have inhabited. The side of the base shows parallel layers of sediment within the seafloor that reflect a lack of burrowing. The top of each base contains a wrinkled surface that is suggestive of the sedimentary structure called “wrinkle structures” that forms in the presence of a seafloor microbial mat. Camptostroma, Helicoplacus, and Lichenoides models designed by Scott Mata in Blender v3.6 and printed on an Elegoo Saturn 3 resin printer. All models are printed using Elegoo ABS-Like Resin except for the arms of Lichenoides that are printed separately using Anycubic Tough Resin for added flexibility.




